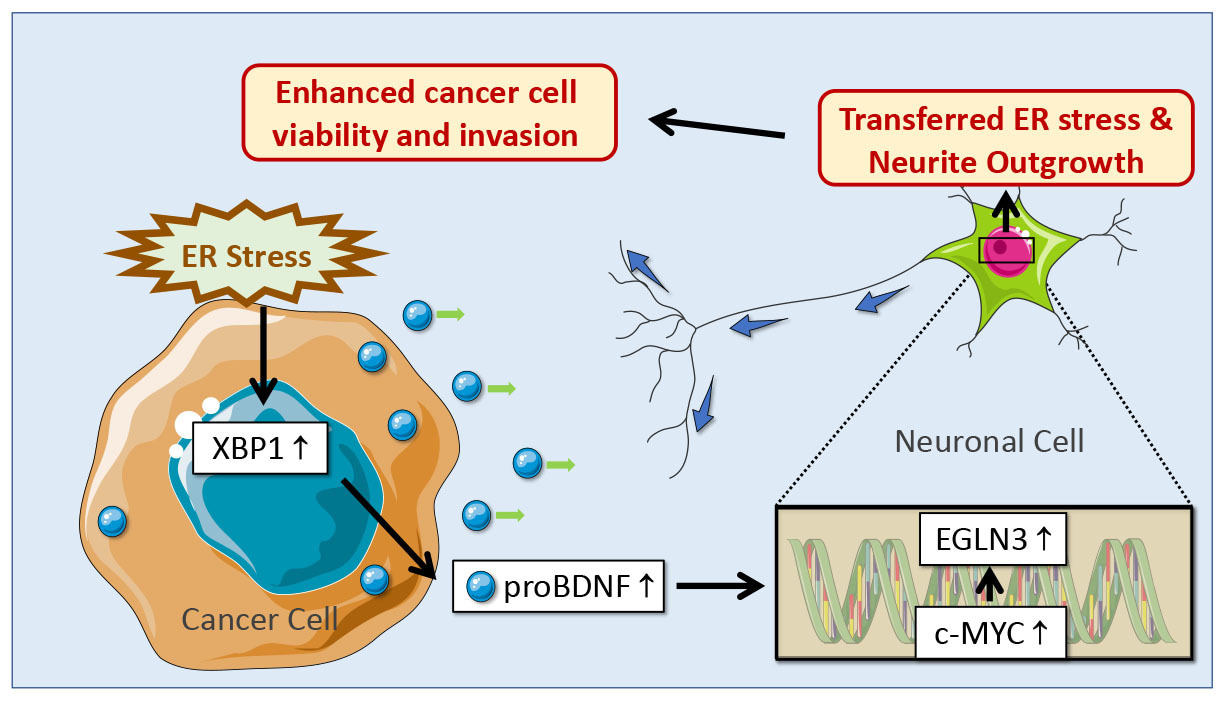
The infiltration of nerves in the tumor microenvironment and the crosstalk between cancer cells and nerves are increasingly described as being essential to tumor growth and dissemination (1, 2). Actionable therapeutic strategies to target nerve development in cancer are emerging (3), but the current impediment is the identification of the initial triggers of tumor innervation. To study the mechanisms initiating the processes leading to tumor innervation, we set our sight on endoplasmic reticulum (ER) stress as it not only significantly contributes to the molecular deregulation leading to tumor growth (4, 5), but is also involved in the development of the nervous system (6-8). This paper was driven by the key question: what is the role of ER stress in tumor innervation?
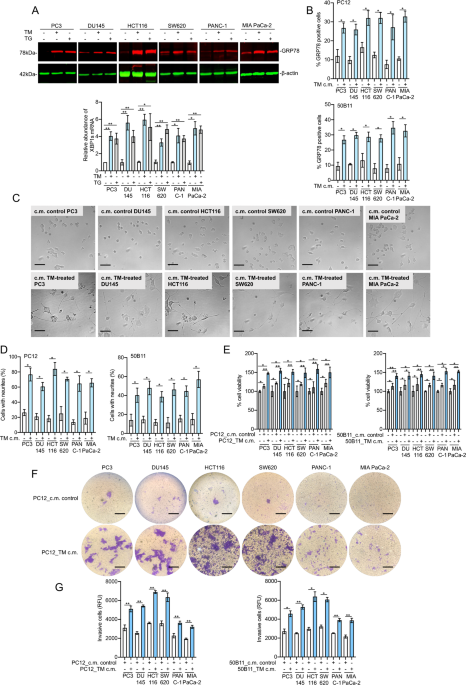
In vitro, we found that conditioned media harvested from ER stressed cancer cells (prostate, colon and pancreatic cancer cells) promoted neurite outgrowth in neuronal cells by transmitting ER stress signal from cancer cells to neuronal cells. To identify the mechanisms responsible for tumor ER stress-induced neurite outgrowth, we screened the release of neurotrophic growth factors by cancer cells (9, 10). ER stress in cancer cells resulted in the secretion of the precursor for brain-derived neurotrophic factor (proBDNF) via the transcription factor X-box binding protein 1 (XBP1), which was necessary for the transmission of ER stress and the promotion of neurite outgrowth. In neuronal cells, next-generation sequencing and mass spectrometry analysis indicated that c-MYC-mediated upregulation of Egl-9 family hypoxia inducible factor 3 (EGLN3) contributed to neurite outgrowth upon ER stress transmission.
After establishing the role of ER stress in promoting nerve outgrowth in vitro (Figure 1), we then validated the finding in vivo by using an orthotopic mouse model of colon cancer. Tumor tissues from mice carrying colon cancer showed the increased expression of XBP1, proBDNF and the growth of nerves upon induction of ER stress. Targeting proBDNF by anti-proBDNF blocking antibody resulted not only in the inhibition of nerve growth in the tumor microenvironment, but also in a decrease in tumor progression. Significantly, 5-Fluorouracil (5-FU), a chemotherapeutic agent used in colon cancer treatment, also promoted colon cancer innervation together with upregulation of the ER stress marker XBP1 and increase of proBDNF.
We finally investigated whether the findings that ER stress in cancer cells induces a XBP1-mediated release of proBDNF leading to tumor innervation could be observed in human tumors. The associations between the expression of XBP1 and that of neuronal markers were found in datasets from The Cancer Genome Atlas (TCGA) for prostate, colon and pancreatic cancers. We then confirm the association between ER stress, proBDNF and tumor innervation by Immunohistochemistry, further consolidating that proBDNF released from ER-stressed cancer cells is driving tumor innervation.
In conclusion, this study reports that ER stress in cancer cells is the trigger of tumor innervation via a mechanism involving the release of proBDNF. This is of clinical significance and suggests that the innervation of the tumour microenvironment is modulated by ER stress-induced by chemotherapy and participates in tumor cell dissemination and cancer relapse. This is particularly important for patients treated with neoadjuvant chemotherapy, where neural sigaling could drive cancer cell dissemination. Our data also show that targeting proBDNF significantly decreased ER stress-induced tumor innervation and reduced cancer growth. Therefore, targeting proBDNF appears to be a possible strategy to limit tumor innervation and enhance chemotherapy outcome.
References
- Faulkner S, Jobling P, March B, Jiang CC, Hondermarck H. Tumor Neurobiology and the War of Nerves in Cancer. Cancer Discov. 2019;9(6):702-10.
- Monje M, Borniger JC, D'Silva NJ, Deneen B, Dirks PB, Fattahi F, et al. Roadmap for the Emerging Field of Cancer Neuroscience. Cell. 2020;181(2):219-22.
- Demir IE, Reyes CM, Alrawashdeh W, Ceyhan GO, Deborde S, Friess H, et al. Clinically Actionable Strategies for Studying Neural Influences in Cancer. Cancer Cell. 2020;38(1):11-4.
- Hetz C, Zhang K, Kaufman RJ. Mechanisms, regulation and functions of the unfolded protein response. Nat Rev Mol Cell Biol. 2020;21(8):421-38.
- Urra H, Dufey E, Avril T, Chevet E, Hetz C. Endoplasmic Reticulum Stress and the Hallmarks of Cancer. Trends Cancer. 2016;2(5):252-62.
- Murao N, Nishitoh H. Role of the unfolded protein response in the development of central nervous system. J Biochem. 2017;162(3):155-62.
- Hayashi A, Kasahara T, Iwamoto K, Ishiwata M, Kametani M, Kakiuchi C, et al. The role of brain-derived neurotrophic factor (BDNF)-induced XBP1 splicing during brain development. J Biol Chem. 2007;282(47):34525-34.
- Valdes P, Mercado G, Vidal RL, Molina C, Parsons G, Court FA, et al. Control of dopaminergic neuron survival by the unfolded protein response transcription factor XBP1. Proc Natl Acad Sci U S A. 2014;111(18):6804-9.
- Allen JK, Armaiz-Pena GN, Nagaraja AS, Sadaoui NC, Ortiz T, Dood R, et al. Sustained Adrenergic Signaling Promotes Intratumoral Innervation through BDNF Induction. Cancer Res. 2018;78(12):3233-42.
- Renz BW, Takahashi R, Tanaka T, Macchini M, Hayakawa Y, Dantes Z, et al. beta2 Adrenergic-Neurotrophin Feedforward Loop Promotes Pancreatic Cancer. Cancer Cell. 2018;33(1):75-90 e7.




Please sign in or register for FREE
If you are a registered user on Research Communities by Springer Nature, please sign in